Return to: MREC Home Page

This predacious mite was accidentally introduced into Germany from Chili in 1958 (Dosse, 1958). From Germany, it was subsequently shipped to other parts of the world, including California (McMurtry et al., 1978) and Florida (Hamlen, 1980). P. persimilis has also become established in southern California (Mcmurtry et al., 1978). It has also been collected in the field in northern-California. According to Kennett and Caltagirone (1968), there are two synonyms for P. persimilis: Phytoseiulus riegeli Dosse and Phytoseiulus tardi (Lombardini).
During the early 1960's, research on this species was conducted in Great Britain, Holland, Canada, and the United States. Since these initial studies, the ability of this predator to control twospotted spider mites has been demonstrated on many plants, including cucumber (Gould, 1970, 1971), tomato (French et al., 1976), ornamental ivy (Gould and Light, 1971), rose (Simmonds, 1972; Boys and Burbutis, 1972), lima bean (Force, 1967), dahlia (Harris, 1971), strawberry (Laing and Huffaker, 1969), and dieffenbachia and schefflera (Hamlen and Linquist, 1981). Although these studies were conducted under greenhouse conditions (or in growth chambers) , there is evidence that P. persimilis can be an effective natural enemy in commercial strawberry plantings (McMurtry et al., 1978) and on ornamentals in commercial interior plantings (Lindquist, 1981). Whether the effectiveness of this predator in these environments will be comparable to its performance under greenhouse conditions remains to be seen.
DEVELOPMENT AND REPRODUCTIVE BIOLOGY
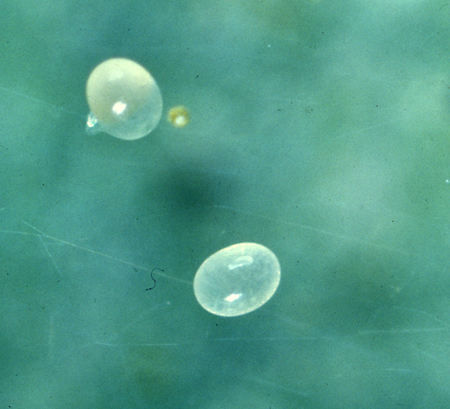
The developmental stage of P. persimilis is similar to that of the twospotted spider mite, ie., egg, larva, protonymph, deutonymph and adult, and has been studied in detail by many authors (Laing, 1968; Sabelis, 1981; Shaw, 1982) . However, the three quiescent periods are absent. The oval eggs are laid in close proximity to a food source. They are light orange and translucent when first deposited, but with age, they darken (Figure 7). The eggs of the predator can be distinguished from those of the prey's by the color as well as the shape (Figure 8).
Click image for larger view. [188kb]
(Figure 7) Eggs of P. persimilis.
Click image for larger view. [178kb]
(Figure 8) Egg of the twospotted spider mite (right) and egg of P. persimilis (left).
Click image for larger view. [127kb]
(Figure 9) Larva of P. persimilis.
The hexapod larva (Figure 9) apparently does not feed and remains inactive unless disturbed. The first feeding stage--the octapod protonymph--emerges from the larval exoskeleton and begins to search for food almost immediately. Feeding and searching continues, with intermittent periods of inactivity presumably due to satiation. The next developmental stage, the octapod deutonymph feeds throughout most of its development. Tho deutonymph later molts, giving rise to the adult; the adult is about the same size as the mature prey mite and is red (Figure 10). Feeding begins soon after molting (Figures 11, 12).
Click image for larger view. [186kb]
Click image for larger view. [158kb]
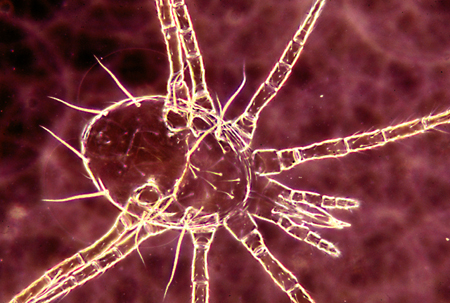
(Figures 10a,b) Adults of P. persimilis.
Click image for larger view. [179kb]
(Figure 10c) Photomicrograph of P. persimilis.
Click image for larger view. [364kb]
(Figure 11) Adult of P. persimilis feeding on an egg of twospotted spider mite.
Click image for larger view. [190kb]
Click image for larger view. [159kb]
(Figures 12a,b) Adults of P. persimilis feeding on immature twospotted spider mites.
Mating usually occurs within a few hours after molting. Multiple matings are common even though the sex ratio is approximately 4 females to 1 male (Laing, 1968). A female that has mated once can lay eggs throughout her life span; whereas an unmated female will not reproduce (Amano and Chant 1978a, 1978b); (Laing, 1968; Schulten et al., 1978). The system of reproduction and sex determination is termed "parahaploidy" (Helle et al.,1978; Hoy, 1982). In this system, both males and females arise from diploid eggs having 8 chromosomes. However, males retain only one complement of 4 chromosomes (haploid) because of the loss or heterochromatization of one half of the chromosomes.
Laing (1968) studied the life history and developed life tables for P. persimilis and T. urticae. His studies were conducted in growth chambers in which the temperature fluctuated between 18-35°C (65-95°F). The time spent in each developmental stage was recorded and various aspects of the reproductive biology were studied. Under these experimental conditions, Laing (1968) determined that P. persimilis would develop from egg to adult in an average of 7.45 days; this is approximately half the time required for development of the twospotted spider mites under similar conditions. For a detailed comparison of its developmental times, at three different temperatures, for various life stages of T. urticae and P. persimilis, see Table 1. Furthermore, the mean generation time (T) was 17.32 days (compared to 24 for the prey), during which the predator population increased 44X (compared to 31X in the prey) (Table 2). Finally, the maximum rate of increase (rmax) for the predator was higher than that for the prey (Table 2). Given these statistics, it is not surprising that P. persimilis is one of the most effective natural enemies of twospotted spider mites known. In fact, at times it can be too effective--i.e., it can often eradicate the prey in the greenhouse (see later section). Table 1.
- *Data obtained on roses in growth chambers (Sabelis, 1981).
**Key to symbols: PN = protonymph, DN = deutonymph, and PO = length of time before an adult female begins to oviposit.The rate of oviposition does not depend on the age of the female, but on the number of eggs previously laid. Eggs will be laid at a rate dependent on conditions until the maximum number is reached or until the female dies from "old age" at about 50 days (Sabelis, 1981). The most important conditions that influence the rate of oviposition are temperature, humidity, and prey density.
Influence of temperature and relative humidity
Temperature has been shown to affect prey consumption, generation time, oviposition, and longevity of P. persimilis (Pruszynski, 1976; Plotnikov and Sadkowskij, 1972; Sabelis, 1981; Shaw, 1982; Laing, 1968; McClanahan, 1968; Takafuji and Chant, 1976). The ultimate outcome of the predator-prey interaction is also influenced by temperature (Force, 1967). The number of deutonymphs eaten by the most voracious stage (the young ovipositioning female) generally increases as the temperature increases. For example at a relative humidity of 75%, the average consumption of spider mite deutonymphs by a single female was 8.8 at 17°C (62.6°F) compared to 13.5 at 26°C (78.8°F) (Pruszynski, 1976). Pruszynski also demonstrated that consumption of prey increased as the relative humidity decreased and the temperature increased. The author also cited a Russian study (Plotnikov and Sadkowskij, 1972) in which spider mite eggs were offered as the prey item. The same trend seemed to occur, i.e., as temperature increased, so did the consumption of eggs. Pruszynski stated that P. persimilis is more sensitive than the prey to temperatures above 30°C (86°F) and that the predator would stop feeding at about 35°C (95°F). The number of eggs and the rate at which they are consumed is also affected by the predator's ability to effectively search for food (see Table 2.)
*Data from Laing (1969).
**Data from Laing (1968).
***Key to symbols: Rmax = Intrinsic rate of increase or the number of individuals produced per female per day. Ro = The number of daughters that replace an average female in. the course of one generation; and T = Mean generation time (days).
The rate at which P. persimilis develops is a function of temperature and is described by a straight line over the range of temperatures between 15-30,C (59-86°F) (Sabelis, 1981); as temperature increases, the time needed to develop decreases (Table 1). However, developmental times in the literature are quite variable and are possibly dependent on the strain studied. Fecundity (number of eggs laid per female) is also influenced by temperature. The temperature at which the maximum reproduction (75 eggs) occurs is approximately 26°C (79°F); the optimum range for reproduction is 17-28°C (63-82°F) (McClanahan, 1968; Sabelis, 1981). At constant temperatures outside this range, females lay fewer eggs. The rate of oviposition, as stated earlier, does not depend on the age of the female, but on the number of eggs previously laid until the maximum number is reached. The effect of temperature on the overall predator-prey interaction was studied by Force (1967). He used constant temperatures of 15°C (59°F), 20°C (68°F), 25°C (77°F), and 30°C (86°F) and obtained excellent control of twospotted spider mites at 20°C; however, at 30°C, P. persimilis was unable to affect control. At 15°C and 25°C, the prey was controlled, but not as dramatically as at 20°C. One problem with this study was the artificial condition of constant temperatures. However, the important points to note are that an optimum temperature apparently does exist and that extreme temperatures can have detrimental effects on the ability of the predator to control the pest population. Despite the temperature limitations, some effort has been made toward developing "heat tolerant" strains (Voroshilov, 1979).
Developmental time can also be affected by relative humidity. A slight increase in the predator's developmental time was observed when the humidity was increased from 40% to 70% (Begljarow, 1967; Ustchekow and Begljarow, 1968; and Stenseth, 1979). Begljarow (1967) noted that development almost stopped at humidities of 25% to 35%. Pralavorio and Almaguel-Rojas (1980) reported that relative humidities below 70% resulted in a significant reduction in the ability of immature predators to molt from one stage to another.
Humidity also exerted an influence on survival of predator eggs at temperatures above 21°C (70°F). At 27°C (81°F) and 40% relative humidity, only 7.5% of the eggs tested in one study hatched compared to 99.7% at the same temperature, but at 80% relative humidity. At 21°C (70°F), there was only a 10% reduction in hatching when eggs were held at the 40% RH compared to those held at 80% (Stenseth 1979). Begljarow (1967) showed that when eggs were held at a relative humidity of 50%, they appeared to shrivel at all temperatures between 13°C (55°F) and 37°C (99°F); while at 60% RH, hatching was successful at temperatures below 30°C (86°F). Sabelis (1981) suggests that the critical relative humidity is 50% and therefore has little influence because the relative humidity in greenhouses rarely falls to levels where predators would be affected significantly.
The searching behavior and activity of P. persimilis can also vary in response to relative humidity. Mori and Chant (1966a, 1966b) investigated the influence of relative humidity on the behavior and activity of this mite and concluded that relative humidity was an important factor limiting the number of prey consumed per predator. In these studies, predator activity and the number of prey consumed per predator increased as relative humidity decreased (i.e., from 100% to 33%). This activity response due to humidity combined with recent evidence (see Sabelis and van der Baan, 1983) that phytoseiid mites, including P. persimilis, are able to use odors (i.e., kairomones) associated with mite infested plants to locate their prey at a distance, further increases the predator's chances of finding and consuming twospotted spider mites.
All developmental stages of the twospotted spider mite are eaten by the adult female P. persimilis. The predator's larval stage does not feed, but the protonymph and deutonymph will feed on the egg, larva, and protonymph stages of spider mites (Takafuji and Chant, 1976). The number of each stage eaten depends on the density of prey and predator, temperature, humidity, stage of predator feeding and which prey stages are available for it to feed upon (Shaw, 1983). Phytoseiulus persimilis depends almost entirely on animals as food (Ashihara et al. 1978, Chant 1961, and Dosse 1958). Ashihara et al. (1978) reported that this predator fed, reproduced, and completed development only on mites in the subfamily Tetranychinae. However, Chant (1961) observed P. persimilis feeding on young thrips. P. persimilis is also cannibalistic when no other food (i.e., spider mites) is available (Dosse, 1958; Laing, 1968). Free- standing water (for the predator to drink) will, in the absence of food, increase survival by 23% (Mori and Chant, 1961b; Ashihara, et al., 1978). Adult females, when fed on honey or a 10% sucrose solution, can survive at least four times longer compared to females being fed on water alone (Ashihara et al., 1978). However, neither sucrose nor free water would promote reproduction. Ashihara et al. (1978) determined that females would not reproduce on a diet of honey, but if they were removed from the honey diet after 35 days and fed spider mite eggs they could achieve their normal reproductive potential.
When compared with five other predatory mites, P. persimilis was rated as having high dispersal powers, and its distribution and that of its prey were highly correlated (McMurtry, 1982). The ability of P. persimilis to disperse and find new colonies of prey depends on the physical characteristics of the environment (Takafuji, 1977), prey distribution and density, predator density, and the duration of infestation or the amount of the spider mite webbing present.
One important environmental characteristic is the density of plants within the greenhouse. For example, when infested plants are dense enough for their leaves to touch, the predator can disperse readily. When the plants have little physical continuity, the predator's ability to disperse can be reduced by about 70% (Takafuji, 1977).
The density of both predator and prey may play a part in the rate at which predators leave an infested plant in search of new sources of food. Young female predators increase the rate at which they depart from a colony as their density increases and that of the prey decreases (Sabelis, 1981; Eveleigh and Chant, 1982). When prey density is low relative to number of predators present, the adult predators begin to disperse in search of new food sources. On the other hand, nymphs of P. persimilis have a much lower capacity--and tendency--to disperse than do the adults and, as a result, they remain behind and feed on whatever food is left before they begin to disperse (Takafuji, 1977). This behavioral characteristic can be a contributing factor to the extinction of prey. Also, the elimination or extinction of the prey in the greenhouse is made possible because P. persimilis has a much greater dispersal potential than its prey (Nachman, 1981). In cases where little or no spider mite damage can be tolerated, such as on ornamental plants, this is a desirable situation. Because some damage can be tolerated in cucumber and tomato crops, it would be desirable to have a stable interaction between the predator and prey over an extended period of time.
The webbing produced by twospotted spider mites aids the searching predator in finding its prey. When webbing is contacted, the predator intensifies its search in the immediate area . The webbing appears to act as an arrestant for dispersing predators. In one study, females were able to find prey twice as fast when webbing was present compared to when webbing was absent (Schmidt, 1976). Schmidt (1976) also reported that spider mite eggs had a similar effect, but to a lesser degree. Kairomones (chemical odors discussed earlier) may be responsible for this nonrandom searching behavior.
The most critical phase in the implementation of any biological control program is the release phase (French et al., 1976; Gould, 1970; Markkula and Tiittanen, 1976). Thus proper timing of predatory mite release is essential to achieve adequate control of the twospotted spider mite. Many of the past failures can be attributed to the detection of natural spider mite infestations too late to utilize biological control successfully (Stenseth, 1980). In these cases, insufficient numbers of predatory mites were released to control an established and rapidly increasing mite population. Consequently, predators were unable to reduce mite numbers fast enough to prevent economic injury.
Release methods have been developed in order to increase the probability of successfully controlling T. urticae with P. persimilis. One release method, termed "Pest-in-First", requires the prey to be released in a uniform pattern before releasing the predator. At the appropriate time, the predator is subsequently released. A specific predator-prey ratio can therefore be established early in the season; the predator also becomes established throughout the greenhouse before mites naturally infest the crop (Markkula and Tiittanen, 1976). A similar technique is the introduction of predator and prey at the same time. This is accomplished by purchasing a mixture of both species from a commercial insectary. In the Netherlands, predators are packed in plastic bottles which contain wheat bran and spider mites. These bottles are sealed with a screw cap equipped with a gauze covered hole for ventilation. This technique, like the "Pest-in-First" method, allows the establishment of specific predator to prey ratios. Thus, balanced control can be attained throughout the greenhouse (Ravensberg et al., 1983).
The rationale behind both of the above techniques is to establish the predator evenly throughout the greenhouse early in the season before the crop becomes naturally infested. This is important where a large portion of the spider mite population enters diapause. Experience has shown that spider mites leave the sites where they diapause and infest the crop as soon as environmental conditions become suitable. Because a large and unpredictable number of mites may enter a crop over a short period of time, severe damage can occur before the problem is noted. These techniques will allow growers the opportunity to establish predators early in the season which will provide a buffer against the immigrating mite population, and thus reduce the potential for damage.
In climates such as Florida, the mass influx of mites, as a result of diapause termination, is seldom seen. Mite infestations occur throughout the year and begin as small isolated patches. Therefore, release techniques designed to release predators when natural infestations are first found are better suited to these conditions (French et al. 1976). Sufficient numbers of predators are released to create a desirable predator-prey ratio, e.g., 1:10 on cucumbers (Markkula and Tiittanen 1976) or 1:6-1:25 on ornamental foliage plants (Hamlen and Lindquist 1981). When distinct patches of prey can be identified, predators should be released on every plant within the patch. Predators should also be released on plants around the outer edge of the infestation in order to establish a barrier that should slow or prevent the spread of the prey. If prey are found, but no distinct patches can be identified, P. persimilis should be released on every fifth plant (Anonymous 1976a - The number of predators to released is dictated more by economics and their availability than any other factor. In situations where the mite population has reached high densities, the cost and logistics for releasing adequate numbers of predatory mites is usually prohibitive. In these cases, conventional chemical control should be employed to reduce the possibility of economic damage.
Many acaricides disrupt the predator-prey interaction even to the extent that acaricides must be applied during the remainder of the growing season. However, studies have shown that this problem can be avoided if either fenbutatin-oxide (Lindquist et al., 1980) or insecticidal soap (Osborne and Petitt, unpublished data) is used as the acaricide. Both studies have demonstrated that a single application of either material used in conjunction with an earlier release of predators gave better control than when either method was used alone.
Once the predators have been released in the greenhouse, some additional conditions should be taken into account. Insecticides, miticides and even certain fungicides can be detrimental to P. persimilis and should be used judiciously. Whenever possible, selective chemicals should be used (see section on Common Problems). As stated earlier, the performance of the predator is conditioned by temperature and relative humidity and to consider this aspect could easily lead to failure of the control program. Sometimes the predator may simply exterminate (eradicate) the prey from the greenhouse; thus, careful monitoring of the mites is required in order to (1) detect such eradication and (2) determine when new introductions of twospotted spider mite occur so that another release of the predator can be made at the proper time.
- Links to other parts of this document
- Introduction
- Twospotted Spider Mite
- Phytoseiulus persimilis (this section)
- Common Problems
- References